INSTITUTO TECNOLÓGICO DE CIUDAD ALTAMIRANO GRO.
LIC. BIOLOGÍA
BITACORA DE LA UNIDAD IV
N.Control:08930129
MATERIA: biologia molecular
Profesor: Francisco Javier Puche Acosta.
Alumna: elidene perez quintana
Vlll semestre
Introducción:
El proceso de replicación de ADN es el mecanismo que permite al ADN duplicarse (es decir, sintetizar una copia idéntica). De esta manera de una molécula de ADN única, se obtienen dos o más "clones" de la primera. Esta duplicación del material genético se produce de acuerdo con un mecanismo semiconservativo, lo que indica que las dos cadenas complementarias del ADN original, al separarse, sirven de molde cada una para la síntesis de una nueva cadena complementaria de la cadena molde, de forma que cada nueva doble hélice contiene una de las cadenas del ADN original. Gracias a la complementacion entre las bases que forman la secuencia de cada una de las cadenas, el ADN tiene la importante propiedad de reproducirse idénticamente, lo que permite que la información genética se transmita de una célula madre a las células hijas y es la base de la herencia del material genético.
La molécula de ADN se abre como una cremallera por ruptura de los puentes de hidrógeno entre las bases complementarias liberándose dos hebras y la ADN polimerasa sintetiza la mitad complementaria añadiendo nucleótidos que se encuentran dispersos en el núcleo. De esta forma, cada nueva molécula es idéntica a la molécula de ADN inicial.
La replicación empieza en puntos determinados: los orígenes de replicación. Las proteínas iniciadoras reconocen secuencias de nucleótidos específicas en esos puntos y facilitan la fijación de otras proteínas que permitirán la separación de las dos hebras de ADN formándose una horquilla de replicación. Un gran número de enzimas y proteínas intervienen en el mecanismo molecular de la replicación, formando el llamado complejo de replicación o replisoma. Estas proteínas y enzimas son homólogas en eucariotas y arqueas, pero difieren en bacterias.
Se conocen cinco polimerasas eucariontes (a, b, g, d y e.) y tienen un mecanismo de acción similar al de las enzimas procariotes.
Las DNA pol. a y d son las que están asociadas mas directamente ala replicación del DNA cromosómico.
La DNA pol. b es responsable de la reparación del DNA. L DNA pol. g se encuentra en las mitocondrias y se encarga de replicar el DNA mitocondrial. La DNA pol. e es recién descubierta y se asemeja en funciones a d.
4.2. Replicación del DNA en eucariontes.
El mecanismo para la síntesis del DNA en eucariontes probablemente es similar al proceso en procariontes.
Las principales complicaciones adicionales son que el DNA en eucariontes se organiza en varios cromosomas lineales y que el DNA es mucho más largo que el que se encuentra en los procariontes. El movimiento de la DNA polimerasa es mucho más lento que en los procariontes; para compensar esto, una célula eucarionte tiene mas o menos 20 000 moléculas de la enzima. Esto permite que se forme un numero mayor de horquillas de replicación, por ejemplo 2000 o más, en los cromosomas eucariontes.
Terminación de la replicación: síntesis de los telómeros
El final de la replicación en eucariotas tiene que resolver el problema del acortamiento de los cromosomas. Para ello ha desarrollado la estructura de los telómeros y telosomas, y la actuación de una nueva enzima emparentada con las retrotranscriptasas: la telomerasa.
|
4.3. Control de la replicación
El proceso resultante de la duplicación de ADN se conoce como división celular, la cual se ha estudiado a nivel citológico estableciéndose ciertas pautas cubiertas por lo que se conoce como ciclo celular.
La mayor parte de los estudios de ciclo celular se han llevado a cabo empleando S. cerevisiae, la levadura del pan y la cerveza también conocida como la levadura de gemación por su característico estilo de división. En el caso de este organismo hay que añadir la ventaja de que actualmente se conocen los aproximadamente 16 millones de pares de nucleótidos que constituyen la secuencia completa de su genoma.
Figura : Ciclo celular de S. cerevisiae
|
|
Estos estudios han permitido identificar una familia de proteínas quinasas dependientes de ciclina, -Cyclin Dependent Kinases, Cdk- que incluyen a Cdc28 de S. cerevisiae, y a su homólogo en S. pombe cdc2, y que juegan un papel central en los procesos claves del ciclo celular: la entrada en ciclo, la síntesis de ADN y la regulación de la mitosis. Estas quinasas son las subunidades catalíticas de un complejo que incluye también na subunidad reguladora, denominada ciclina, necesaria para la función del complejo al determinar la localización o la especificidad de sustrato de la quinasa.
4.4.1 Secuenciación del ADN
Secuenciacion de los Acidos Nucleicos:
La introducción de métodos rápidos de secuenciación a finales de los años 70 representó un cambio importante en los estudios sobre el DNA Existen TRES métodos de secuenciación de los ácidos nucleicos. Uno de ellos (Maxam y Gilbert) utiliza reactivos químicos a fin de cortar el DNA a nivel de bases específicas.
Método enzimático de terminación de cadena o método didesoxi de Sanger:
Para obtener la secuencia de bases nitrogenadas de un segmento de ADN por el método enzimático de terminación de cadena, se necesitan los siguientes compuestos:
El ADN molde o segmento de ADN que se desea secuenciar. Para poder secuenciar un segmento de ADN, previamente se necesita tener gran cantidad de ese fragmento, y por tanto, hay que clonarlo en un vector apropiado. Además, debe estar en estado de hélice sencilla.
Breve descripción del método automático de secuenciación
La principal diferencia entre método enzimático de terminación de cadena y el método automático de secuenciación radica, en primer lugar en el tipo de marcaje. En el método automático en vez de radiactividad se utiliza fluorescencia y lo habitual es realizar cuatro mezclas de reacción, cada una con nucleótido trifosfato (dTTP) marcado con un fluorocromo distinto. Este sistema permite automatizar el proceso de manera que es posible leer al mismo tiempo los ADNs de nueva síntesis producto de las cuatro mezclas de reacción.
Objetivos:
_ El alumno entenderá la replicación del ADN.
Metodologia:
En esta unidad abarcaremos con el tema de la replicación ya que es el es el mecanismo que permite al ADN duplicarse es decir, sintetizar una copia idéntica también hablaremos del experimento de Meselson y Stahl que permitió demostrar que el mecanismo real se ajusta a la hipótesis de replicación semiconservadora. Este blogger publicaremos los temas vistos en clases asi como también las prácticas.
UNIDAD 4 REPLICACIÓN DEL ADN:
La propiedad más notable de las células vivas es su capacidad de reproducirse con una fidelidad casi perfecta, no solamente una generación, sino durante centenares y millares de generaciones. El modelo que explica cómo se duplica el DNA se denomina modelo semiconservativo ya que a partir de una molécula de ADN obtendremos dos exactamente iguales, cada una con una hebra antigua (cadena parda en la imagen siguiente) y otra nueva (cadena naranja en la imagen siguiente): la cadena original comienza a separarse de un extremo a otro de modo que cada una de las hélices va a sintetizar una hélice nueva complementaria habiendo tomado como modelo o patrón a la hélice inicial.

El mecanismo de replicación es es esencialmente el mismo en todas las células. Es un proceso semiconservativo porque cada uno de los dos ADN hijos tiene una cadena del ADN anterior.La replicación del DNA está catalizada por una enzima que se denomina DNA-polimerasa: cataliza la formación de cadenas de nucleótidos por la adición sucesiva de nucleótidos trifosfato (NTP).
Al incorporarse el NTP, se liberan dos de los tres fosfatos para aportar la energía necesaria a la reacción química. Las DNA-polimerasas sólo funcionan utilizando un DNA como molde, necesitando inexcusablemente un cebador (pequeño oligonucleótido de 5 a 30 nucleótidos que puede ser un pequeño DNA o RNA) al que se añadirá el primer nucleótido. Los nucleótidos se van añadiendo en el extremo 3’ de la cadena en síntesis.
Un complejo sistema de regulación intracelular le indica el momento del ciclo celular en que debe comenzarse la replicación, la enzima reconoce una secuencia de nucleótidos llamada origen de replicación que marca el punto el que ha de comenzarse la replicación. En Escherichia coli es la secuencia de 245 pares de bases que tiene varias repeticiones de una secuencia consenso GATCTNTTNTTT.
Meselson y Stahl confirman en 1958 que la síntesis del DNA es semiconservativa partiendo de un DNA con gran contenido de [15N].
El proceso se inicia con la enzima helicasa que se dedica a deshacer los puentes de hidrógeno entre los pares de bases para poder separar las dos cadenas del DNA.
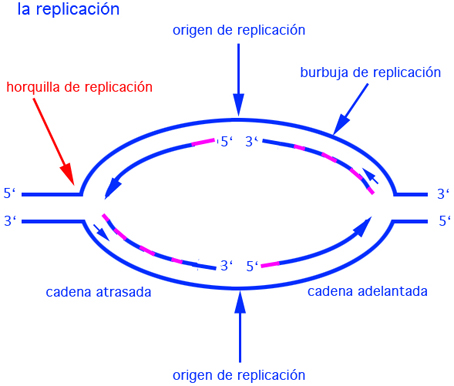
En el caso de E. coli, se observa que el origen de replicación es rico en A y T para que cueste menos energía separar las cadenas. La separación de las dos cadenas provoca una serie de tensiones topológicas en el DNA que hacen necesaria la intervención de otras enzimas, las topoisomerasas, para eliminar dichas tensiones.Se forma así una burbuja de replicación con un ADN desapareado flanquedado por sendos ADN apareados.
Una vez que se abre la molécula, se forma una área conocida como "burbuja de replicación" en ella se encuentran las "horquillas de replicación" . Por acción de la la ADN polimerasa los nuevos nucleótidos entran en la horquilla y se enlazan con el nucleótido correspondiente de la cadena de origen (A con T, C con G). Los procariotas abren una sola burbuja de replicación, mientras que los eucariotas múltiples. El ADN se replica en toda su longitud por confluencia de las "burbujas".
Durante este proceso, la ADN polimerasa reconoce una base nucleotídica no apareada de la cadena original y la combina con un nucleótido libre que tiene la base complementaria correcta. Luego, la ADN polimerasa cataliza la formación de nuevos enlaces covalentes que ligan el fosfato del nucleótido libre entrante con el azúcar del nucleótido previamente agregado en la cadena hija en crecimiento. De esta forma, la ADN polimerasa sintetiza el esqueleto de azúcar-fosfato de la cadena hija.
Dado que las cadenas del ADN son antiparalelas, y que la replicación procede solo en la dirección 5' to 3' en ambas cadenas, numerosos experimentos han mostrado que, una cadena formará una copia continua, mientras que en la otra se formarán una serie de fragmentos cortos conocidos como fragmentos de Okazaki . La cadena que se sintetiza de manera continua se conoce como cadena guía, delantera ó adelantada y, la que se sintetiza en fragmentos, cadena atrasada ó rezagada.

Los fragmentos cortos, recién sintetizados de hebras de ADN se denominan fragmentos Okazaki. Todas las ADN polimerasas conocidas, pueden sólo sintetizar ADN en la dirección 5' a 3' . Sin embargo, cuADNo las hebras se separan, la horquilla de replicación se desplaza a lo largo de una hebra molde en la posición 3' a 5' y 5' a 3' en el otro lado del molde.
En la primera , la cadena adelantada de ADN es sintetizada continuamente en la dirección 5' a 3'. En la otra, llamada la cadena atrasada, la síntesis de ADN sólo ocurre cuADNo una sección de una hebra simple de ADN ha sido expuesta y procede en la dirección opuesta a la dirección de la horquilla de replicación (5' to 3'). Es por ésto discontinua y la serie de fragmentos Okazaki son unidos de manera covalente por las ligasas formADNo una hebra continua. Se denominan fragmentos Okazaki porque fué éste investigador quien primeramente los observó y estudió usADNo timidina radioactiva . En los eucariotas, los fragmentos Okazaki están formados por unos cuantos cientos de nucleótidos, mientras que en los procariotas estos fragmentos pueden alcanzar algunos miles.
Para que la ADN polimerasa trabaje, se necesaria la presencia, en el inicio de cada nuevo fragmento, de pequeñas unidades de ARN conocidas como cebadores, a posteriori, cuADNo la polimerasa toca el extremo 5' de un cebador, se activan otras enzimas, que remueven los fragmentos de ARN, colocan nucleótidos de ADN en su lugar y una ADN ligasa que los une a la cadena en crecimiento.
El conjunto de proteínas que se forman el punto de bifurcación de la horquilla se conoce como replisoma.
Una visión general del proceso es la siguiente:

La biosíntesis de la hebra guía y rezagada esta coordinada, por que la hebra rezagada forma un bucle de manera que físicamente apunta en la misma dirección que la hebra guía. Esto significa que ambas hebras tiene un acceso similar a la enzima polimerasa.
Estructuras en q
La replicación del DNA circular produce las llamadas estructura en q que se forma como resultado de las dos horquillas de replicación. La replicación se da simultáneamente en la dirección de las manecillas del reloj y en la dirección contraria. Para exponer las hebras sencillas de DNA, las dos hebras progenitoras deben girar. Por cada 10 bases que se copian el DNA debe dar 1 vuelta completa. Puede calcularse que en E. coli la horquilla de replicación necesita moverse a a una velocidad de 800 bases por segundo, lo que requiere que el DNA progenitor se desenrolle a razón de 80 revoluciones por segundo.

El desenrollamiento del DNA induce a un super-enrollamiento negativo (en la dirección opuesta). La replicación semiconservativa del DNA requiere que se produzca un corte en uno o en ambos nucleótidos del esqueleto para aliviar la tensión en la molécula, esta labor la hace las topoisomerasas. La topoisomerasa I cambia el DNA superenrrollado a un DNA relajado, La topoisomerasa II (girasas) produce hélices negativas al destorcer el DNA
4.1 Replicación del genoma procariótico
Aunque las actividades enzimáticas sean las mismas en procariotas y eucariotas, hay muchas más proteínas implicadas en la replicación de los eucariotas. Por otro lado, el DNA eucariótico tiene más problemas con el superenrollamiento al estar muy estrechamente asociado a unas proteínas denominadas histonas. También es distinto el hecho de que en un cromosoma procariótico sólo hay un origen de replicación en el único cromosoma, mientras que en los de eucariotas suele haber muchos más (a veces más de 30.000) por cada cromosoma.
DNA polimerasas procariotas
Las DNA polimerasas I, II y III se encuentran en las procariontes. En la siguiente tabla se resumen las propiedades de las diferentes enzimas.
Propiedades de la DNA polimerasa de E: coli y de retrovirus
La DNA polimerasa I de E. coli es una sola cadena polipeptídica de Mr 109 000. Contiene un átomo de zinc por molécula. Cataliza la polimerización en el sentido 5´ 3´.
Todas las polimerasas de procariotes muestran actividad exonucleasa, es decir pueden hidrolizar DNA, en procariontes pueden hidrolizar DNA de cadena sencilla (en la dirección 3´ 5´) y de cadena doble (en la dirección 5´ 3´).
LA DNA polimerasa III de E. coli esta formada por 7 subunidades: a, b, g, d, e, q y t. Las siete son escenciales para la actividad máxima.
La DNA pol. III es la principal enzima de replicación en E. coli. La DNA pol. I es responsable de la reparación de DNA dañado. La funcion de la DNA pol II no esta todavía clara.
4.2. Replicación del DNA en eucariontes.
El mecanismo para la síntesis del DNA en eucariontes probablemente es similar al proceso en procariontes.
Las principales complicaciones adicionales son que el DNA en eucariontes se organiza en varios cromosomas lineales y que el DNA es mucho más largo que el que se encuentra en los procariontes.
Terminación de la replicación: síntesis de los telómeros
El final de la replicación en eucariotas tiene que resolver el problema del acortamiento de los cromosomas. Para ello ha desarrollado la estructura de los telómeros y telosomas, y la actuación de una nueva enzima emparentada con las retrotranscriptasas: la telomerasa.
4.3. Control de la replicación
El proceso resultante de la duplicación de ADN se conoce como división celular, la cual se ha estudiado a nivel citológico estableciéndose ciertas pautas cubiertas por lo que se conoce como ciclo celular.
La mayor parte de los estudios de ciclo celular se han llevado a cabo empleando S. cerevisiae, la levadura del pan y la cerveza también conocida como la levadura de gemación por su característico estilo de división.
Polimerasa
|
E. coli
|
|
|
Virus*
|
|
I
|
II
|
III
|
|
Mr
|
109 000
|
120 000
|
180 000
|
160 000
|
Estructura, subunidades, Mr
|
una
|
una
|
a 140 000
|
a 65 000
|
|
|
|
e 25 000
|
b 95 000
|
|
|
|
q 10 000
|
|
Polimerización 5´ 3´
|
+
|
+
|
+
|
+
|
Actividad exonucleasa
|
|
|
|
|
5´ 3´
|
+
|
-
|
+
|
-
|
3´ 5´
|
+
|
+
|
+
|
-
|
virus de la mieloblastosis avícola, un retrovirus